Introduction
Human biology has many alarm systems. Some are obvious: fever, pain, inflammation, tachycardia and fatigue. Others are molecular and silent. Growth differentiation factor 15, commonly abbreviated as GDF-15, belongs to this second category. It is a circulating stress-responsive cytokine and hormone-like protein that rises when cells, tissues and organs are under pressure1–3. For many years, GDF-15 was mainly considered a biomarker: a blood signal associated with ageing4,5, cardiovascular disease6–8, cancer9–11, kidney disease12, inflammation13,14, mitochondrial dysfunction15,16 and poor prognosis. More recent research has changed this view. GDF-15 is not merely a passive marker of disease. It can actively communicate with the brainstem, reshape appetite, influence energy balance, contribute to nausea, regulate tissue tolerance during inflammation, participate in cancer cachexia and modulate immune responses.
This makes GDF-15 one of the most fascinating molecules in modern translational medicine. It sits at the crossroads of stress biology, metabolism, inflammation, pregnancy, oncology, cardiology and ageing. In simple terms, GDF-15 is a biochemical message that says17–20:
The body is under stress; conserve energy, change behaviour and adapt physiology.
In disease, however, that same adaptive signal may become harmful, especially when chronically elevated
What is GDF-15?
GDF-15 is a divergent member of the transforming growth factor-β superfamily. It was first described in the late 1990s as macrophage inhibitory cytokine-1 (MIC-1),or NSAID-activated gene 1 (NAG-1) because early experiments showed that it could inhibit lipopolysaccharide-induced tumour necrosis factor-α production by macrophages1. Later, it became widely known as GDF-15.
Unlike many classical cytokines that mainly act locally, GDF-15 circulates in the blood and can function as an endocrine stress signal2,17,18. Under healthy basal conditions, circulating concentrations are generally low, although they increase with age4. During physiological or pathological stress, levels may rise substantially. GDF-15 is induced by mitochondrial dysfunction15,16,21, endoplasmic reticulum stress21,22, hypoxia23, oxidative stress24, DNA damage25,26 inflammation13,14, tissue injury27,28, pregnancy29–31 and malignant disease9–11. A useful way to understand GDF-15 is to think of it as a cellular distress hormone. When cells experience danger or metabolic strain, they release GDF-15 into the circulation. The body then interprets this signal, particularly through the brainstem, and changes appetite, nausea, energy handling and immune-neuroendocrine responses17,29,32.
The GDF-15–GFRAL axis: how the body sends distress signals to the brain
For many years, a major question remained unanswered: through which receptor does GDF-15 produce its biological effects? The breakthrough came in 2017, when several independent groups identified GFRAL or glial cell line-derived neurotrophic factor family receptor alpha-like, as the high-affinity receptor for GDF-1517–20. GFRAL is expressed mainly in the Area Postrema (AP) and Nucleus Tractus Solitarius (NTS) of the brainstem.
GDF-15–GFRAL Axis
Cellular stress
↓
GDF-15 released into blood
↓
GDF-15 reaches brainstem
↓
GDF-15 binds GFRAL
↓
GFRAL recruits RET
↓
RET becomes activated/phosphorylated
↓
Brainstem neurons are activated
↓
Reduced appetite, nausea, food aversion, weight loss, autonomic and immune-neuroendocrine effects
This anatomical location is biologically elegant. The area postrema is one of the brain’s circumventricular organs, meaning it has a relatively permeable blood–brain barrier. It can sample circulating signals and trigger nausea, vomiting, appetite suppression and aversive responses17,33,34. Therefore, GDF-15 is not simply a peripheral inflammatory marker. It is a blood-borne signal that can reach a specialised brainstem detection system.
When GDF-15 binds GFRAL, the receptor complex recruits the co-receptor RET, activating downstream neuronal signalling17–20
- RET stands for REarranged during Transfection.
It is a receptor tyrosine kinase, meaning it is a cell-surface signalling protein that can activate intracellular pathways when stimulated.
This pathway suppresses food intake, promotes weight loss and may generate nausea or food aversion depending on context33,34. In animal models, loss of GFRAL prevents many of the anorectic and weight-reducing effects of GDF-1517–20. These findings transformed GDF-15 biology from a descriptive biomarker field into a mechanistic neuroendocrine discipline.
GDF-15 at the Interface of Physiology and Disease
Although GDF-15 is frequently discussed in the context of disease, it also has important physiological roles in stress adaptation, metabolic regulation and maternal–fetal biology. Figure 1 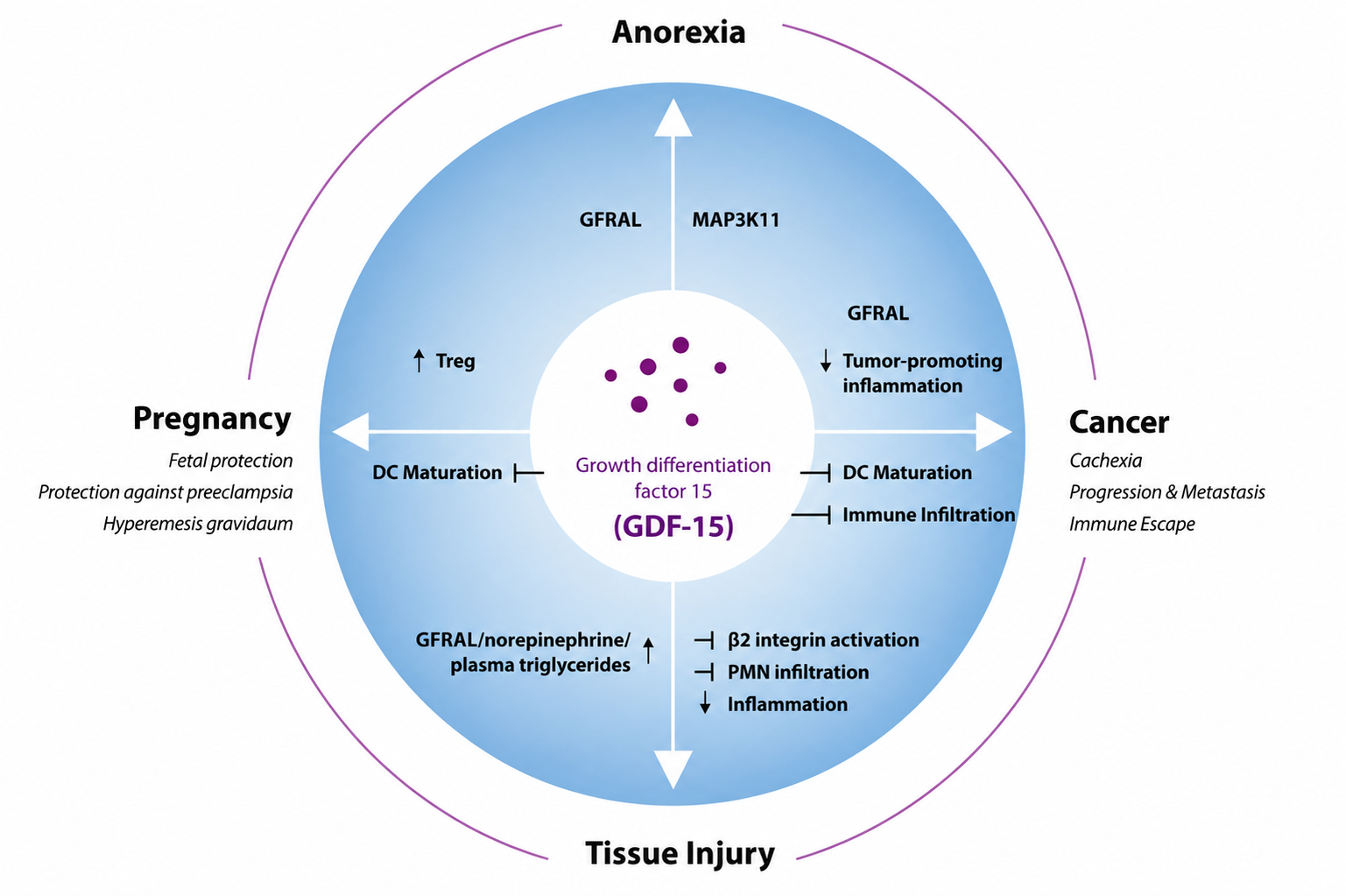 Figure 1: GDF-15 in physiological and pathological contexts. provides a concise overview of these key physiological and pathological contexts.
Figure 1: GDF-15 in physiological and pathological contexts. provides a concise overview of these key physiological and pathological contexts.
Figure 1: GDF-15 stimulates or inhibits different cellular processes via GFRAL and potentially other still unknown receptors in various physiological and pathophysiological situations. DC, Dendritic cells; Treg, regulatory T cells35.
Ageing and baseline variation
Circulating GDF-15 increases with age. Large population studies show that older adults generally have higher concentrations than younger adults4. This likely reflects cumulative cellular stress, low-grade inflammation, mitochondrial strain, vascular injury, renal function changes and multimorbidity. Therefore, GDF-15 is sometimes described as a biomarker of biological ageing or “inflammageing”4,5,12.
Inflammageing
Inflammageing refers to the chronic, low-grade, systemic inflammatory state that develops with advancing age. It is not usually driven by an acute infection, but by cumulative biological stress, including cellular senescence, mitochondrial dysfunction, oxidative damage, impaired autophagy, immunosenescence, adipose-tissue inflammation, gut-barrier changes and persistent release of damage-associated molecular patterns.
This produces a sustained rise in inflammatory mediators such as IL-6, TNF-α, IL-1β and C-reactive protein. Over time, inflammageing contributes to frailty, atherosclerosis, insulin resistance, neurodegeneration, cancer vulnerability and reduced resilience to physiological stress.
However, interpretation requires caution. A high GDF-15 level is not diagnostic of one disease . It is more like smoke from a fire: it tells us something is burning, but not exactly what is burning. Age, smoking, diabetes, cardiovascular disease, kidney function, pregnancy and malignancy can all influence circulating concentrations2,4.
Mitochondrial stress
Mitochondria are the cell’s major energy-producing organelles31. When mitochondrial function is impaired, cells activate integrated stress responses, including transcriptional programmes that increase GDF-15. This is why GDF-15 has emerged as a useful biomarker in mitochondrial disorders. In several clinical studies, patients with mitochondrial myopathies or mitochondrial respiratory chain disorders had elevated serum GDF-15 compared with controls or patients with other myopathies. This does not mean GDF-15 alone can diagnose mitochondrial disease. Rather, it may help clinicians decide who needs deeper investigation, such as genetic testing, muscle biopsy or specialised metabolic evaluation15,16.
Pregnancy
Pregnancy is one of the most striking physiological states in which GDF-15 rises. The placenta produces high levels of GDF-15, especially early in pregnancy. This has become highly relevant to understanding nausea and vomiting of pregnancy and hyperemesis gravidarum, discussed later. From a physiological perspective, the placenta may use GDF-15 as part of a maternal–fetal signalling system. From a clinical perspective, excessive maternal sensitivity or sudden exposure to high GDF-15 may contribute to severe pregnancy sickness29,30.
Metformin and metabolic adaptation
Metformin, one of the most widely used medicines for type 2 diabetes,increases circulating GDF-15. Experimental and clinical studies suggest that part of metformin’s effect on body weight and energy balance may be mediated through GDF-15. This does not mean all metformin benefits are GDF-15-dependent, but it shows that a classic diabetes drug may influence a brainstem-linked stress hormone pathway36,37
GDF-15 in appetite,nausea and body weight regulation
GDF-15 is now recognised as a central regulator of illness-associated anorexia11,17,38.During infection, cancer, organ failure, chemotherapy exposure or systemic inflammation, appetite often declines. This response may be adaptive in the short term because it redirects energy away from digestion and toward survival38. However, when prolonged, it can contribute to malnutrition, sarcopenia and cachexia9,11.
The GDF-15–GFRAL pathway appears to act less like a traditional appetite hormone such as leptin or ghrelin and more like a danger-associated feeding brake17,33,34,38. It is activated in response to injury, toxins and cellular stress rather than ordinary meal-to-meal energy balance. This distinction matters because GDF-15 does not simply tell the brain, “energy stores are sufficient.” It may tell the brain, “the body is ill; eating may be unsafe or unnecessary right now”17,38.
This biology helps explain why GDF-15 is attractive but complicated as a therapeutic target. Activating the pathway could reduce appetite and body weight, which may be useful in obesity Blocking the pathway could improve appetite and weight in cancer cachexia11,17,20.But the same pathway is also linked to nausea and aversive symptoms. Therefore, the therapeutic window must be carefully defined29,33,34.
GDF-15 and hyperemesis gravidarum
One of the most important recent developments in GDF-15 biology comes from pregnancy research. Nausea and vomiting are common in early pregnancy, but hyperemesis gravidarum is a severe disorder characterised by persistent vomiting, dehydration, weight loss, electrolyte abnormalities and, in severe cases, hospitalisation29,30. Historically, hyperemesis gravidarum was poorly understood and, at times, wrongly psychologised. The emerging GDF-15 literature has helped shift the field toward a mechanistic biological explanation29,31,39.
The first major genetic signal came from a genome-wide association study showing that variants near GDF15 and IGFBP7, genes involved in placentation, appetite regulation and cachexia biology, were associated with hyperemesis gravidarum30. This genetic evidence was supported by prospective pregnancy data showing that higher circulating GDF-15 concentrations around week 15 of gestation were associated with vomiting and antiemetic use during pregnancy39. A subsequent serum biomarker study found that GDF-15 and IGFBP7 levels were significantly increased in women hospitalised with hyperemesis gravidarum at 12 weeks’ gestation, whereas hCG levels were not significantly different between cases and controls40. These findings helped weaken the older assumption that hCG alone explains severe pregnancy sickness.
Further support came from whole-exome sequencing, which identified additional GDF15 variants associated with hyperemesis gravidarum and strengthened the argument that GDF-15 is not simply correlated with symptoms but may contribute to disease susceptibility41. More recently, a Nature study showed that fetal and placental production of GDF-15, together with maternal sensitivity to the hormone, contributes substantially to nausea and vomiting in pregnancy and hyperemesis gravidarum29. A large 2026 multi-ancestry Nature Genetics study further confirmed GDF15, GFRAL, IGFBP7 and other loci involved in appetite, insulin signalling and brain plasticity as part of the genetic architecture of severe pregnancy nausea and vomiting31.
The key concept is therefore not simply “high GDF-15 equals hyperemesis.” A more nuanced model is emerging: the placenta produces large amounts of GDF-15, but maternal susceptibility may depend on pre-pregnancy exposure, genetic background and sensitivity of the GDF-15–GFRAL brainstem pathway. This helps explain why many pregnancies have high GDF-15, but only some develop severe, disabling vomiting.
The clinical implications are still developing. Future strategies may include risk prediction, targeted biomarker testing, desensitisation approaches or modulation of GDF-15 signalling. However, any intervention in pregnancy must be evaluated with exceptional caution because GDF-15 may also have protective roles in maternal–fetal biology .
GDF-15 in cardiovascular disease
Cardiovascular medicine was one of the first fields to recognise the prognostic value of GDF-15. GDF-15 rises in myocardial stress, ischemia–reperfusion injury, heart failure, endothelial dysfunction, oxidative stress and systemic inflammation2,6–8,42,43. Experimental work suggested cardioprotective roles in ischemia–reperfusion injury, while clinical studies showed that circulating GDF-15 predicts adverse outcomes6–8,42,43.
In patients with acute coronary syndromes, high GDF-15 has been associated with increased risk of major bleeding, myocardial infarction, stroke and mortality6,7. In atrial fibrillation and coronary artery disease, it has been incorporated into biomarker-based risk frameworks2,6,7. In heart failure, GDF-15 provides prognostic information beyond traditional clinical variables, reflecting a broader burden of biological stress8.
But GDF-15 is not a cardiac-specific biomarker like troponin. Troponin indicates myocardial injury. Natriuretic peptides indicate myocardial wall stress. GDF-15 is broader: it reflects systemic stress, inflammation, ageing, kidney function, metabolic disease and vascular injury. Its power lies in risk stratification, not disease specificity2,4,8.
For clinicians, the key message is this: GDF-15 may help identify vulnerable patients, but it must be interpreted in context.
GDF-15 in cancer
Cancer is one of the most clinically important areas of GDF-15 research. Many tumours produce GDF-15, and high circulating levels often correlate with advanced disease, systemic inflammation, weight loss and poor prognosis. The role of GDF-15 in cancer is complex and context-dependent. In some early settings, stress-induced GDF-15 may participate in growth control or tissue protection. In advanced malignancy, however, tumours may exploit GDF-15 to support immune evasion, anorexia and cachexia9–11.
Cancer cachexia
Cancer cachexia is a devastating syndrome characterised by involuntary weight loss, skeletal muscle wasting, anorexia, fatigue and metabolic dysfunction. It is not simply starvation. Patients with cachexia lose muscle and fat because inflammatory and tumour-derived signals alter metabolism, appetite and tissue turnover9–11.
GDF-15 is now recognised as a major driver of cancer-associated anorexia and weight loss in some patients. By activating GFRAL-positive neurons in the brainstem, tumour-derived GDF-15 can suppress appetite and contribute to weight loss17–19. This mechanism has moved rapidly from biology to clinical trials.
A major advance came with ponsegromab, a humanised monoclonal antibody that inhibits GDF-1511. In a phase 2 trial published in the New England Journal of Medicine, ponsegromab improved body weight, appetite-related symptoms and physical activity in patients with cancer cachexia and elevated GDF-15. This was an important proof-of-concept: blocking a circulating stress hormone could improve a clinically meaningful wasting syndrome11.
Immune evasion and immunotherapy resistance
Another major frontier is tumour immunology. Recent studies indicate that tumour-derived GDF-15 can impair T-cell recruitment into the tumour microenvironment Figure 2 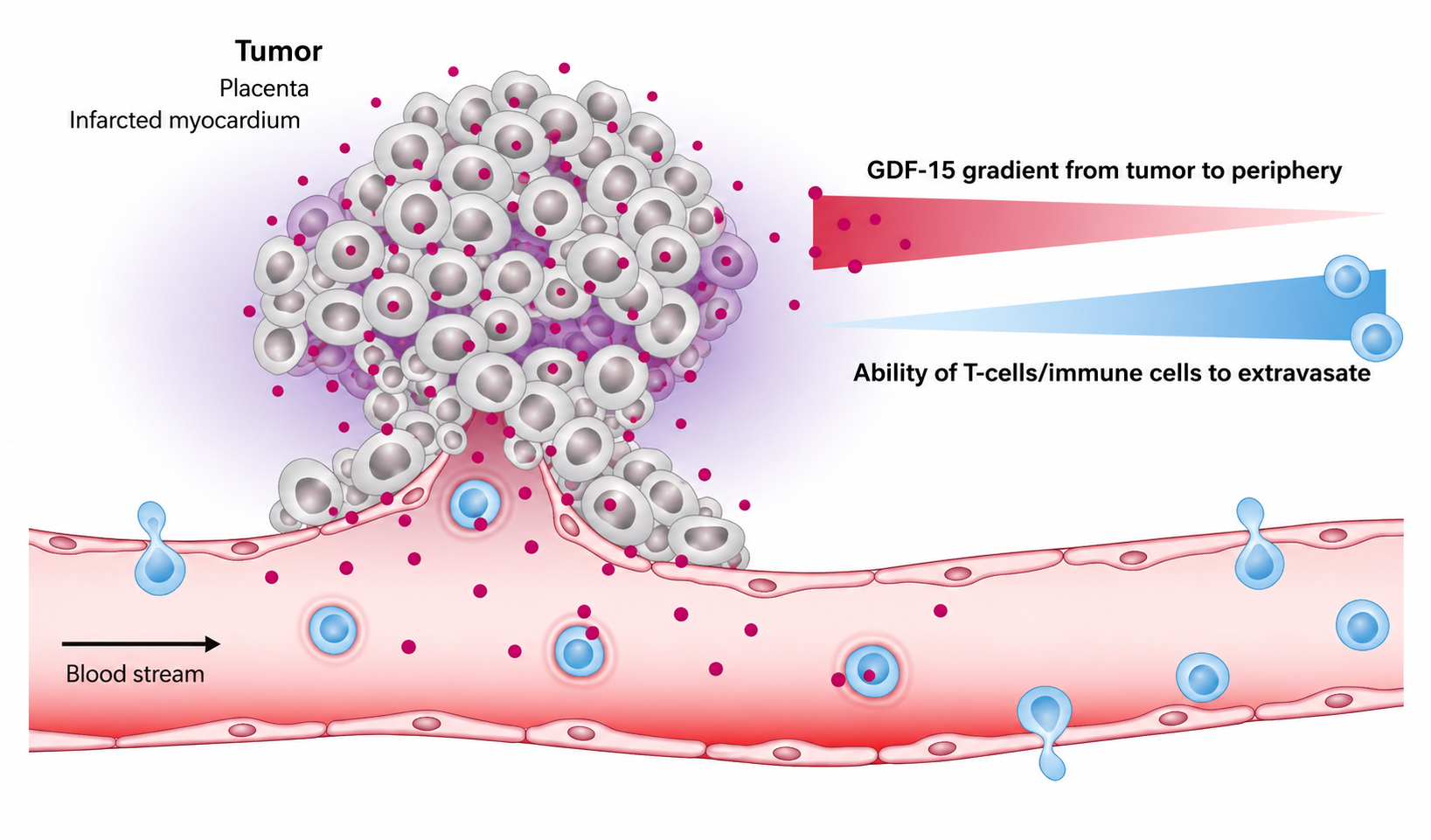 Figure 2: The role of GDF-15 in immune modulation and T-cell infiltration. 9,10. One mechanism involves disruption of LFA-1-dependent T-cell adhesion and migration. In simple terms, GDF-15 may act like a molecular “keep out” signal that prevents immune cells from entering tumour tissue effectively9.
Figure 2: The role of GDF-15 in immune modulation and T-cell infiltration. 9,10. One mechanism involves disruption of LFA-1-dependent T-cell adhesion and migration. In simple terms, GDF-15 may act like a molecular “keep out” signal that prevents immune cells from entering tumour tissue effectively9.
Figure 2:The role of GDF-15 in immune modulation. In various pathological conditions, GDF-15 correlates inversely with the ability of T cell to infiltrate the tumor, placenta, or the infarcted myocardium35.
This has therapeutic implications. Neutralising GDF-15 may help overcome resistance to immune checkpoint inhibitors such as anti-PD-1 and anti-PD-L1 therapies9,10. Recent Nature research on visugromab, a GDF-15-blocking antibody, suggests that GDF-15 neutralisation may restore immune infiltration and improve responses in some patients with refractory cancer10. These findings remain early but highly significant because resistance to immunotherapy is one of the central challenges in oncology.
GDF-15 in inflammation and tissue tolerance
Inflammation is not only about killing pathogens. The body must also survive the damage caused by infection and immune activation. This concept is called disease tolerance or tissue tolerance Figure 3 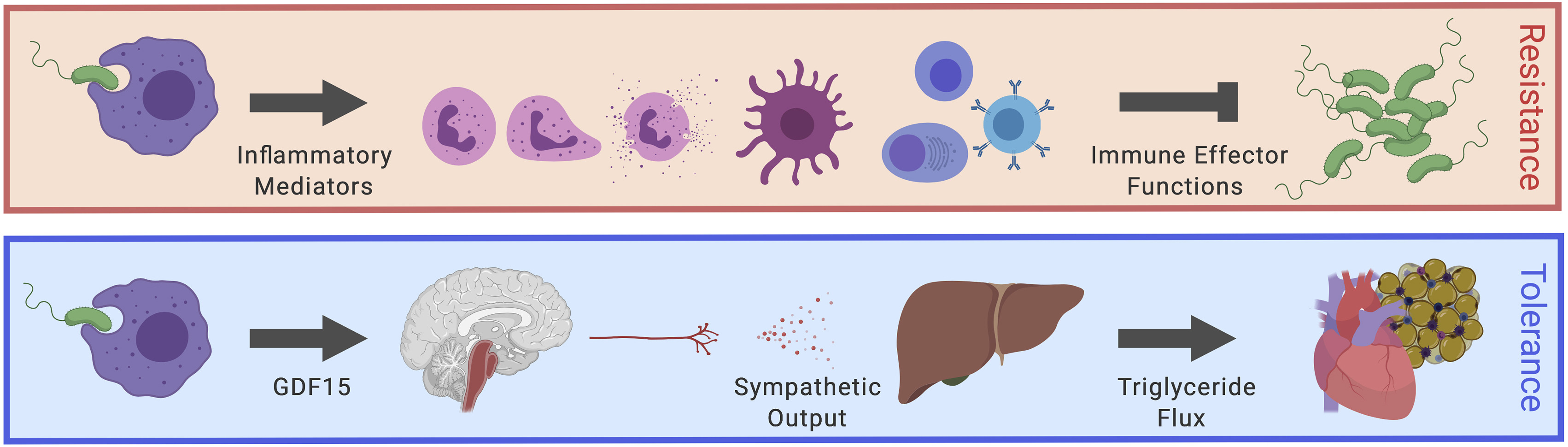 Figure 3: Model of GDF15 regulating hepatic sympathetic outflow and lipid metabolism. . It refers to the ability of the host to limit tissue injury without necessarily reducing pathogen burden44,45.
Figure 3: Model of GDF15 regulating hepatic sympathetic outflow and lipid metabolism. . It refers to the ability of the host to limit tissue injury without necessarily reducing pathogen burden44,45.
Figure 3:Model of GDF15 regulating hepatic sympathetic outflow controlling Lipid metabolism to support cardiac function38
Resistance
Bacteria or virus enters the body
↓
Immune system detects the pathogen
↓
Neutrophils, macrophages, antibodies and complement are activated
↓
Pathogen burden decreases
↓
Infection is controlled
Tolerance
Infection causes inflammation
↓
GDF-15 increases
↓
GDF-15 supports hepatic sympathetic signalling
↓
Blood triglyceride levels are maintained
↓
Heart function and body temperature are protected
↓
Survival improves despite inflammatory stress
A landmark Cell study showed that GDF-15 is induced during inflammation and can coordinate tissue tolerance through metabolic regulation, particularly triglyceride metabolism38. In experimental models, GDF-15 was important for survival during bacterial and viral inflammatory stress. This shifts our understanding of GDF-15 from “bad marker of disease” to “adaptive survival molecule”38.
However, biology is rarely one-dimensional. A molecule that protects during acute inflammation may become harmful when persistently elevated. In chronic disease, continuous GDF-15 signalling may contribute to anorexia, frailty, immunosuppression or cachexia9–11,38. Thus, the same pathway can be protective in acute stress and maladaptive in chronic disease.
GDF-15 and autoimmunity: The brain–spleen–T-cell axis
A particularly exciting recent development is the discovery that GDF-15 may regulate immune responses through the nervous system32. Sonner et al in 2026 reported that a GDF-15–GFRAL axis can suppress autoimmune T-cell responses during neuroinflammation. In experimental autoimmune encephalomyelitis, a mouse model used to study multiple sclerosis-like neuroinflammation, GDF-15 accumulated in the inflamed central nervous system, and its absence impaired inflammation resolution32. The proposed pathway involves GDF-15 activation of GFRAL-positive neurons in the brainstem, followed by sympathetic signalling to the spleen, increased norepinephrine synthesis and reduced expression of integrins required for autoreactive T-cell trafficking32.
This finding is important because it places GDF-15 within the broader field of neuroimmune regulation. In simple terms, GDF-15 may act as a circulating stress signal that tells the brainstem to reduce harmful immune-cell movement into vulnerable tissues32. This does not mean GDF-15 is universally anti-inflammatory. Rather, it suggests that, under certain conditions, GDF-15 may function as a physiological brake on excessive immune-mediated tissue injury32,38.
Several additional studies support the broader immunological relevance of GDF-15. A landmark Cell study showed that GDF-15 is induced during inflammation and coordinates tissue tolerance, allowing the host to survive inflammatory stress without necessarily suppressing pathogen clearance38. Clinical studies in rheumatoid arthritis found that serum GDF-15 levels were higher in patients than controls and correlated with disease activity, morning stiffness, tender joint count, erythrocyte sedimentation rate and vascular changes46. More recent data in primary Sjögren’s syndrome showed that serum GDF-15 was significantly elevated in patients, associated with extra-glandular manifestations and correlated with disease activity47. In autoimmune uveitis, experimental work showed that activation of GDF15 downstream of the transcription factor EGR2 could modulate retinal microglial phenotype and alleviate autoimmune ocular inflammation48.
Together, these studies suggest that GDF-15 is not merely a general inflammatory biomarker. It may participate in active immune regulation across several disease contexts, including neuroinflammation, rheumatic disease and autoimmune eye disease. However, the field is still young. In some settings, elevated GDF-15 may reflect tissue injury or disease severity rather than direct immune protection. Therefore, the most balanced interpretation is that GDF-15 is a context-dependent immunometabolic signal: protective during some forms of acute or resolving inflammation, but potentially maladaptive when chronically elevated or exploited by disease processes.
GDF-15 as a biomarker: useful, but not disease-specific
Because GDF-15 rises in many conditions,it is best viewed as a systemic stress biomarker2,4,5.It has potential clinical value in cardiovascular risk stratification2,heart failure prognosis8, acute coronary syndrome bleeding and mortality risk6,7, mitochondrial disease screening15,16, cancer cachexia phenotyping11, frailty and biological ageing assessment4,5, pregnancy nausea and hyperemesis gravidarum research29, inflammatory and infectious disease prognosis13,38.
Yet the same breadth creates a limitation. GDF-15 is not specific. A raised level does not tell us whether the problem is cancer, heart failure, kidney disease, mitochondrial dysfunction, pregnancy, inflammation or ageing2,4. It must be interpreted alongside clinical history, examination, standard laboratory tests, imaging and disease-specific biomarkers. In future medicine, GDF-15 may be most useful as part of multi-biomarker panels rather than a standalone diagnostic test.
Therapeutic possibilities: should we block or activate GDF-15?
The therapeutic question is fascinating because both directions may be useful in different diseases.
Blocking9–11
Blocking GDF-15 may be helpful when the hormone is driving harmful appetite suppression, cachexia or tumour immune evasion.
Cancer cachexia is the clearest example.Ponsegromab and visugromab represent a new generation of therapies designed to neutralise GDF-15.These approaches could potentially improve weight, appetite, activity and treatment tolerance in selected cancer patients.
GDF-15 blockade may also enhance immunotherapy responses by allowing better T-cell entry into tumours. This area is still developing, but it is one of the most promising oncology applications.
Activating17–20,36,37,49
Activating the GDF-15–GFRAL pathway may reduce appetite and body weight, making it theoretically attractive for obesity and metabolic disease. However, nausea and aversion are major concerns and the challenge is to separate them from beneficial metabolic effects.
Modulating sensitivity29
Pregnancy research introduces a third possibility: modifying sensitivity to GDF-15 rather than simply blocking or activating the pathway. If low baseline exposure increases susceptibility to hyperemesis gravidarum, carefully timed pre-exposure or desensitisation strategies might one day be considered. This remains experimental and must be approached cautiously.
The paradox of GDF-15: protector, biomarker and disease driver
The central lesson of GDF-15 biology is paradox. GDF-15 can be protective during acute stress but harmful during chronic elevation. It can signal tissue damage, but also coordinate adaptation. It may reduce inflammation and improve tissue tolerance, but tumours may exploit it to avoid immune attack. It may help the body survive infection, but incancer it may contribute to anorexia and wasting. It may explain pregnancy nausea, but it may also participate in normal placental communication. This duality is not unusual in biology. Cortisol, interleukin-6, TGF-β and nitric oxide all have protective and harmful effects depending on timing, dose, location and disease context. GDF-15 belongs to this family of context-dependent molecules.1,2,10,11,38
Conclusion
GDF-15 has emerged from relative biological obscurity as a centralintegrative signal linking cellular stress, mitochondrial dysfunction, inflammation, metabolic adaptation, neuroendocrine regulation and immunemodulation. Its relevance extends across pregnancy biology, cardiovascular disease, cancer cachexia, ageing and autoimmune neuroinflammation, illustrating how a single stress-responsive cytokine can participate in diverse physiological and pathological networks. Importantly, GDF-15 is no longer best understood as a passive biomarker of disease severity; accumulating evidence indicates that it can act as a mechanistic mediator of appetite regulation, tissue tolerance, immune-cell trafficking and systemic adaptation to injury.
For students and early-career scientists, GDF-15 provides a powerful example of how biomarker biology can evolve into disease-mechanism biology. For clinicians, it represents a promising indicator of systemic vulnerability, biological stress and prognostic risk, although its lack of disease specificity requires careful interpretation within the broader clinical context. For researchers and drug developers, the GDF-15–GFRAL axis offers an increasingly important therapeutic frontier, particularly in cancer cachexia, immunotherapy resistance, hyperemesis gravidarum, metabolic disease and neuroimmune regulation. The future clinical value of GDF-15 will depend on precision: defining when its elevation is adaptive, when it is maladaptive and when targeted modulation may improve human health.